[拼音]:wuwei mu
[外文]:Salientia,Anura
两栖纲中种类最多,体形最特殊的1目。成体无尾和鳃。现有20余科300余属3500余种。除南极洲外,广布于各大洲;在温差小,湿度大的中美、南美和非洲热带和亚热带种类最多,个别种达北极圈南缘。在全北界有3个大科只有模式属:蟾蜍科(共约有25属340种)中只有蟾蜍属,雨蛙科(共约有40属600余种)中只有雨蛙属,蛙科(共约有50属670种)中只有蛙属(Rana),而且这些地区的物种也少。在古北界以外的亚洲和非洲,蟾蜍科和蛙科的其他属种占有优势;在美洲,蛙科只有蛙属,蟾蜍科有少数其他的属;在热带美洲,雨蛙科占有绝对优势,属种极多。在热带非洲和亚洲,虽然没有雨蛙属,但有在生态、形态方面与雨蛙科趋同适应的非洲树蛙科和树蛙科(共约有20余属400种)。此外在中美、南美,还有一个占绝对优势的大科即细趾蟾科(共约有50属700余种) 和特有的小科(共约有5科12属近200种)。在大洋洲没有蟾蜍属和蛙属种类;雨蛙类及细趾蟾类分别被另列为浜雨蛙科(共有4属近150种)及龟蟾科(共有20属近百种)。其他主要科的分布区:盘舌蟾科(见铃蟾属)仅分布于旧大陆;锄足蟾科(近10属80余种)的锄足蟾亚科分布于欧洲、北非和北美,属种少(2~3属10余种);角蟾亚科分布于亚洲东部和东南部,中国属种多(50种以上);异舌蟾科是一个单型科属种,主要分布于墨西哥。呈断裂分布跨越两洲者有滑蹠蟾科(2属4种)分布于大洋洲和北美,另外有负子蟾科(4属26种)分布于南美和非洲。姬蛙科广泛分布于环球温带至热带的一个大科,属种甚多(共约有60余属近280种)。亚洲没有特有科。
中国已知近230种,分隶7科25属。角蟾亚科的属种在中国最多,髭蟾属、齿蟾属等为中国的特有属。无尾类中约有 2/3的种主要分布于横断山脉和青藏高原地区。
形态与结构体宽而短,后肢长,跗部自成一节,适于跳跃或游泳(图1)。皮肤一般光滑湿润,有的皮肤上有角质刺,或头顶部皮肤骨质化而全无小鳞,皮肤与皮下肌肉之间有一些大淋巴囊。口大,舌后端多游离,可翻出摄食。下眼睑连有透明的瞬膜。眼大,位于头侧,突出时可扩大视野。下颌无齿,上颌一般有细齿。中耳多完备,鼓膜显著或隐于皮下或无。雄蛙一般有共鸣器(声囊),这是一种第二性征(图1)。额骨与顶骨愈合为额顶骨,有方轭骨,副蝶骨⊥形(图2)。椎体有双凹型、后凹型、变凹型、间凹型或参差型(图3)。椎骨一般为10枚,荐椎后为一细长的尾杆骨腰带呈“U”字形,髂骨长,平直向前与荐椎横突相关联。脊柱短而数少,弯曲度和灵活性不强;前三对躯椎上或有短肋,是原始性状,见于低等类群。脊柱与后肢的关联由髂骨、后肢骨和尾杆骨部位的肌肉、肌腱而加强,可以作有力的攀援、跳跃或对称蛙式游泳动作。前肢较强,主要可减轻落地时的冲击力。肩带左右侧上喙骨或相接(固胸型)或相互重叠(弧胸型)(图4),或有弧固型。桡骨与尺骨、胫骨与腓骨分别愈合为桡尺骨和胫腓骨;近端 2跗骨长,自成1节,增强了跳跃、游泳能力。无尾类体形体现了与运动行为相适应的结构。多数种类大多“蹲在以后肢向后掘的浅泥凹内,既可防失水,又能坐待捕食,捕食时后肢仍贴于地面。




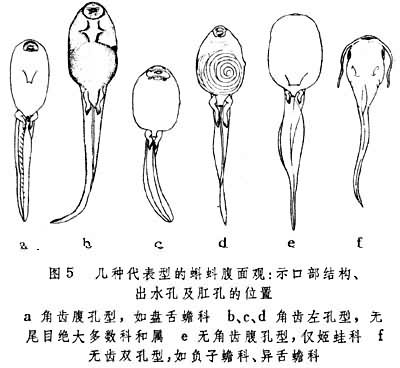
无交接器,体外受精。卵生,仅个别种卵胎生或胎生。幼体──蝌蚪的体形、食性等与成性迥然不同。蝌蚪口部或有角质小齿;有鳃和尾,早期为外鳃,不久外鳃萎缩,代之以内鳃;舌弓部位皮肤逐渐向后生长,成为覆盖着内鳃的鳃盖褶,再与腹位的皮肤愈合,形成鳃盖腔,仅留有1或2个小出水孔,出水孔或位腹面中部或两侧或位体左侧,出水孔通向体外,依据角齿之有无和出水孔之特征,分为4个类型:
(1)角齿腹孔型(图5a);
(2)角齿左孔型(图5b、c、d);
(3)无角齿腹孔型(图5e);
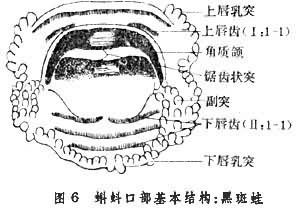
(4)无角齿双孔型(图5f)。口部一般有成行的角质小齿和角质颌(图6)。以刮取藻类为主,无者以吞食浮游生物为主。蝌蚪先出后肢芽,前肢于早期就在鳃盖腔内或附近部位发育,变态高潮时,前肢伸出体外,尾、鳃萎缩以至消失,器官改组,呼吸和消化器官尤为突出。此时一般不摄食,也不能久潜于水中,否则大多要溺死。
 生活习性
生活习性
本目动物有水栖、半水栖、陆栖、树栖、穴居等不同的种类。一般栖息于各种淡水水域及其附近潮湿的林木草丛中。在黄昏、黎明或雨后的白天活动频繁,觅食各类昆虫及其幼虫和蠕虫等。繁殖习性多样化,有以各种方式抚育卵或幼体的种类,如著名的负子蟾,幼体在雄蛙声囊中的尖吻达蛙、小蛙在胃内的胃育溪蟾、雨蛙科体背有各式抚育结构,如在背面皮肤表内的囊蛙等。此外,有的可在卵内发育成蝌蚪直至完成变态,然后小蛙破卵膜而出,这种直接发育方式(蝌蚪期不在水中)在热带潮湿地区的无尾类中常见。蝌蚪期有的长达3年;性成熟期短的数月,长的3年以上。一般在冬眠出蛰后抱对产卵,在湿热地区,有的种类可常年产卵。
起源与系统发生马达加斯加发现的早三叠世化石三叠尾蛙,曾称原蛙,有与近代型无尾目基本相似的主要特征。一般认为它是无尾类的先驱,形态上已具备向典型无尾类演化发展的基础;如头骨与肢骨基本形态,脊柱趋短,骼骨延长;有胁骨及尾椎骨示原始性状。
近代型的有在阿根廷发现的比较完整的侏罗纪化石,如侏罗南蛙等;在以色列曾发现有许多完整的早白垩世化石无舌类(Aglossa=负子蟾类);此外,还有锄足蟾类的蝌蚪和近于细趾蟾类的化石。在侏罗纪末期至白垩纪早期,几个主要原始类群都已出现,而且分化速度相当快。一般认为滑蹠蟾科(双凹型)、盘舌蟾科和负子蟾科(后凹型)的个体发育时成体或亚成体期有3~4对肋骨,属于原始类群,被列为始两栖亚目,还包括没有肋骨异舌蟾科(后凹型)和锄足蟾科(变凹型);其余的10余科属于高级类群,被列为新两栖亚目。锄足蟾科被认为是介于原始型的和高级型的中间类群。