[ƴ��]��dengshua ranseti
[����]��lampbrush chromosome
�����ˢ״����һ�ദ����չ״̬��������ת¼�Ļ�״ͻ��ľ�Ⱦɫ���������ڽ��м������ѵ�ϸ���С����������ͬԴȾɫ������γɵĺ��� 4��Ⱦɫ����Ķ����塣��ĸϸ�������������ȫ��mRNA���������ʶ��Ǵӵ�ˢȾɫ��ת¼�����ϳɵġ�
��ˢȾɫ���״���W.��������1882�걨������δ�϶�����һ��Ⱦɫ�塣1892��J.����������������ĸϸ���е����־�Ⱦɫ��Ľṹ�������о�����������״����ŷ��ʱ��ϴú�͵��ֵĵ�ˢ��ȡ��Ϊ��ˢȾɫ�塣50�����H.G.�������˸Ľ����о�������ˢȾɫ����о������չ������
��ˢȾɫ���������ֲ��㷺�������ֵ�ʱ�ڲ�ͬ�����������ĵ�ˢȾɫ���������ĸϸ����һ�μ������ѵ�˫���ڣ��������嶯���֫�����еļ���ʹ���������ࡢë���Ƥ�����ͼ��������Щ�����ˢȾɫ������ھ�ĸϸ���������ѵ�ǰ�ڣ��������ֱ��Ŀ��˫��Ŀ���棬�����������Լ�⨺���˵����鶯�����߳�ĵ�ˢȾɫ��ɳ����ڼ�����ϸ���С���ֲ�����ijЩ���͵�ϸ�����ຬ�е�ˢȾɫ�塣�ߵ�ֲ���������Ӽ���������Ҳ���ֵ�ˢȾɫ�壬���紹���к����ס�
��ͬ���ֵ�ˢȾɫ��Ĵ�Сʮ�����⡣һ��˵����������Ⱦɫ����DNA�ĺ�����Cֵ���ߵĶ����ˢȾɫ���������ҵ�ˢȾɫ�����ʱ��Ҳ�Ƚϳ���������ľ�����ߵ�Cֵ����ˢȾɫ��Ҳ������о���ˢȾɫ��ij��ò��ϣ������о�����Ϊ�꾡����ijЩ��β�����ࡣ
�������ˢȾɫ��
��������ĸϸ���У���һ�μ������ѵ�˫����ʱ��ÿ��Ⱦɫ�������ͬԴȾɫ�徭���ƶ��γ�����Ϊ4�����Ⱦɫ����Ľṹ����������(ͼ1)����һ״̬����ĸϸ���п�ά�����»�����֮�á�

ÿ��Ⱦɫ������һ��ȥ�����Ǻ˵��� (DNP)��ά���ɡ���ά��һ����λ��װ���ܳ�ΪȾɫ��������ɺ̵ܶ�DNP��������Ϊ��˿��Ⱦɫ������˿���ɵ�ˢȾɫ����ᡣ��Ⱦɫ��������չ����DNP��ά��Ϊ���ͼ2�������ϳ���⣬������˿�������塢������������Ƚṹ����Щ�ṹ��ijЩ�����������ض�Ⱦɫ��Ĺ̶���λ���ݴ˿��Լ������Ⱦɫ�壬��Ϊ��ꡣ
 Ⱦɫ��
Ⱦɫ��
���ͬԴȾɫ���У�Ⱦɫ������Ⱦɫ������Ŀ�ͷֲ�������ͬ������״���ܹ���������Ⱦɫ����ɨ��羵�¿ɼ�������С�Ͳ��ɡ�Ⱦɫ���������϶࣬������ĵ�����Ⱦɫ������Լ��5000��Ⱦɫ����Ⱦɫ������Ŀ�ʹ�С�����������йأ�Ҳ��ת¼���Զ��仯���������ĸϸ�������ڲת¼���Դ��ڸ߷壬Ⱦɫ����С������Ŀ�϶ࡣ��ʱ�������߾���D���Ʋ��ת¼���ԣ����ڵ�Ⱦɫ����˴��ں϶���Ⱦɫ�����ϲ��������ϴ�������塣�����������������ĸϸ���������죬�ת¼�������ͣ��������Ⱦɫ��Ҳ�˴����ںϣ��������٣����ճ�һ�����ϳ�Ⱦɫ����Ⱦɫ���ᡣ
��˿����ˢȾɫ�����˿��ͨ��λ�ڽ��渽������˿���ж�����̬��һ��Ϊ����״����С��̬��ǰ��Ⱦɫ���������֣��������������һ������˿����ǰ�����������Ⱦɫ���˴��ں϶��ɵ�����������ͼ3������˿��������Ͼ������ˢȾɫ����˿��ָ������ϸ���Ĵ����ͬ��
 ���ת¼
���ת¼
���DNA��Ծ��ת¼���������ǵij������ܴ��������вƽ������Ϊ10��15�ף����Ŀɴ�200��300�ף�С���ڹ�ѧ���������Էֱ档ͨ��һ��Ⱦɫ����һ�Բ������ЩȾɫ��������������ڲ�ϴ�ת¼��ʼ�㵽�յ��Ϊһ��ת¼��λ���õ羵��չ���۲쵽�DNAϸ˿��ת¼����:ֱ��ԼΪ12.5��һϵ�е�RNA�ۺ�ø������ת¼��λ��DNAϸ˿�ϡ�ת¼������Ǻ˵���(RNP)����ʼ���϶̣��Ժ�������������ת¼��λ�Ŀ�խ��ͬ������Լ2��3�ף���ʹÿһת¼��λ����̬�ϳ��ּ��ԡ��������ֻ��һ��ת¼��λ����Щ��������ת¼��λ����Щ��λ��ת¼���������ͬ��Ҳ���Բ�ͬ��ͼ4��������ת¼����(����-RNP)�����ࡢ�����Ͷѻ�״̬�IJ�ͬ����ijЩȾɫ������ض���λ�����γɲ�ͬ���͵IJ��ͼ5������Щ��ǵ�ˢȾɫ�����Ҫ��ꡣ

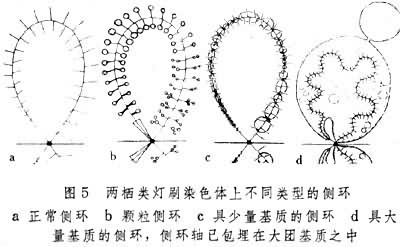
���������ͬ���ز����ġ�ijЩ��ܳ����ڲ�ͬ����̶ȵ���ĸϸ���ĵ�ˢȾɫ���ϣ�����һЩ�����������ĸϸ������չ����ʾ��Ծ��ת¼���ԣ��Ժ�ʧȥת¼���Բ�������Ⱦɫ���С��ڽϴ����ĸϸ���г��ֽϳ��IJ������Ҳ�١����ش���Ҳ��Ӱ������չ�ͻ��������������������������͵Ľṹ����Щ��̬�仯�������в������
�������ض���λ���ֵ�����(ͼ6)�൱��һ��Ⱦɫ��Ĵ��˺ۣ����������Ե��ס����������Ե����䵽�����У�����ͬһ��λ�γ��µ����塣����䳣���˴��ں�������С�������ӡ����������¿ɼ������ڲ�ΪԲ�����ܵ����ı������ܶȽ��������ʰ�Χ��(ͼ7)�� �ڲ�ͬ������֮������Ĵ�С����Ժ㶨�ġ�

 ����
����
λ��ijЩȾɫ�����ˣ���һ���ڵ������а�����DNP��С���塣��������������С���ֵ����ԡ�
������ṹ��������ơ�Ҳ�ɼ������������ںϡ���Щ�������Ⱦɫ���Ͼ����Զ��Ϻ㶨��λ�ã�����Ϊ����Ⱦɫ��Ľ�ꡣ
����������ͼ����ijЩ�������ˢȾɫ����������������꣬��˫��ṹ�����״�ṹ�����Խϴ�IJ�ɴصس���������ض���λ�ȡ�����̬ѧ�����������Ѿ�����˶����������ˢȾɫ��Ĺ���ͼ��(ͼ8����������ͼ��ӳһ����ˢȾɫ�����Գ��ȡ���˿����λ���ֲ�ͬ���������λ�ã������õ�ˢȾɫ����������о��Ļ�����

��Ӭ��YȾɫ��
���ֶ��ᆱĸϸ��Ⱦɫ���Ͽ��Կ�����Χ����ë״�Ľṹ����ʾ���ӷ����д��ڵ�ˢ�ڡ����ֹ�Ӭ�������ڵij�����ĸϸ���еĵ�ˢ���ṹ�Ƚϵ��ͣ��о���Ҳ��Ϊ�����
��Ӭ��ĸϸ����һ�γ�����ѵ�˫���ڣ���ijһȾɫ���ϻ���ֳɶԵIJ������XO�͵Ĺ�Ӭ�ľ�ĸϸ���в��߲����XYY��Ӭ�ľ�ĸϸ���в��Ŀ�ӱ��� ����������ж�ֻ��YȾɫ���ǵ�ˢȾɫ�塣YȾɫ���ڼ��������в���ԣ����Ժ�������IJ�ͬ����Ӭ�ĵ�ˢȾɫ�岻�Ƕ����塣��Ӭ�� YȾɫ�干��ƽ������Ϊ50�� ��5�Բ(ͼ9)��ÿ�Բ����ר�ŵ����ơ�
 ����
����
λ��Ⱦɫ��̱��ϵ�һ�Բ��DNP�Ậ260ǧ�����(kb)���ڳ�����Ƭ�Ͽɼ�������ֱ��Ϊ40����RNP�������ǵ��͵ĵ�ˢ�����
��״��λ�ڳ��۽���˿����������2������ܴ����״���������״����0.2��1�ĵ������������ɢ����˿������ɡ���Բ�Ƿ����ˢ���������ת¼��Ŀǰ��ȱ��Ⱦɫ����չʵ���֤�ݡ�
��״����Բ����ѧ�����ɼ��ɺ�����С��������ɢ������ɡ��ڵ��������¿ɼ���ɢ������ֱ��35����С����ɣ������ܵĿ�����С�ܽ��ܰ�װ���ɡ���չʵ�������״���Ľṹ���Ƶ�ˢ���
�ٺ�����Բ���д�ļٺ��ʺ�ǰ���һ���ٺ��������߳��ṹ���ƣ�������˿������ɣ����к���С�� RNP��������չʵ���пɼ�ת¼������1000kb��˵����Բ�Ͽ���ֻ��һ��ת¼��λ��
˿״��λ�ڳ��۶˲�������������ɡ�Զ�˵���ɢ��������˿��ɣ����зֲ���35��40���ĸ���RNP�Ŀ�����һ����ٺ�������ӣ����˵����ܲ����ɽ��ܰ�װ����˿��ɣ���RNP��������չʵ��֤��DNP�����г�Լ500kb��ת¼��λ��
��Ӭ��ˢȾɫ��IJ��Ծ�غϳ�����Ⱦɫ�岻�ܺϳɵ�һ�ֶ������ز����ٵ�RNA��û��YȾɫ���XO���۹�Ӭ����ȫ�����ġ� YȾɫ����ȱ�ݵĹ�Ӭ���ӷ����г��ֵ���̬ѧ�쳣��ʮ�ֶ����ģ�����ϸ�����쳤ʱ�������裻������ϸ������ֱ������Լ���Щϸ�������������쳣�����˵����ϵĻ���Ļ��ܣ���YȾɫ�������������ӣ�����Ϊ��Щϸ�����ֻ�ʱ�����ijһ�����ʱ��룬���ǵ�����ϸ�������;���ϸ����������װ�䡣��������YȾɫ����εĹ�Ӭ���侫�ӷ������������ڻ����ڣ�ȡ����ȱ����һ�Բ������ʾ��ͬ���������ӿ������ε������ھ����γɵIJ�ͬ�ĽΡ�
ֲ��ĵ�ˢȾɫ��
�й�ֲ���ˢȾɫ��ı������ࡣ�ڴ��������Լ�������˫���ڡ��������Լ��������ձ��ھ��е�ˢ�ڵļ��ء����������ĵ�ˢȾɫ�壬ֻ��һ���ϳ���Ⱦɫ�壬��Χ����ë״�Ľṹ��ͼ10���������������˫���ڵĵ�ˢȾɫ����˴�ͬС�졣ֻ�����к�ɡ����е��͵ĵ�ˢȾɫ��Ľṹ��ͼ11����


��ˢȾɫ����һ���װ����һ��Ⱦɫ����ô���ܣ��������ڽ���ת¼�ľ�Ⱦɫ�壬����������о�Ⱦɫ��ṹ���ܵĶ��ز��ϡ���ˢȾɫ���о�����ȡ�õĽ�չ��Ҳ�ٽ��˶�һ��Ⱦɫ��ṹ���˽⡣
70���������������ͬλ�ر�ǵ�DNA���ˢȾɫ�����²����� RNA����ԭλ�ӽ�����ȷ����������Ķ�λ������½��������ЩDNA̽��רһ����ǿ�������鵰�����5sDNA̽�붼����������ת¼��RNA�ӽ�������ˢȾɫ��ͬһ����Ҳ��Ϊ��ͬ̽������ǣ�������ʹ���Ƕ������ַ�������λ�Ļ������ʵ�Բ����˻��ɡ�
���Ÿ���ȷ��ʵ�鼼���ķ�չ��Ӧ�ã�����Ԥ�������ӷ����е�ˢȾɫ��Ļ������õ��������ⲻ������������̥���ڷ���ʮ����Ҫ�����һ��������˽���ϸ���ֻ��л���ĵ�������ơ�