[ƴ��]��hesuan
[����]��nucleic acid
һ�������������ʣ����Ŵ���Ϣ��Я���ߡ������������ӽṹ�����ı����������л�ܵ����ţ��������Ŵ�����������ı��졣���������ϸ�����з��֣��Ҷ��Ծ����ԣ���������֪�������һ�ж�ֲ��ϸ���Լ���������ϸ���Ļ�����ɳɷ֡�������ϸ���ṹ�ĸ��ֲ������ಡ������Ҫ��Ψһ��ɳɷ֡�������ˣ���������������������������ֳ���Ŵ���������ش��������������������á�
���ݻ�ѧ��ɵ��ص㣬����ɷ�Ϊ���Ǻ�����RNA�����������Ǻ��ᣨDNA��������;ǰ�߷����ж���D-���ǣ����߷����ж���D-2-�������ǣ����ߵķ��ӽṹ������ѧ�������IJ���ͬ������RNA��DNA��ֻ������ϸ��֮�ڣ���ϸ�����ʻ�ϸ����Һ��������ڡ����ݹ��ܺͷֲ��IJ�ͬ��RNA�ֿɷ�Ϊת�˺��Ǻ���(tRNA)����ʹ���Ǻ���(mRNA)�ͺ˵�������Ǻ���(rRNA)���࣬���Ǿ��ֲ���ϸ���ʵIJ�ͬ��ϸ���ṹ��������������ϳ��и������Ŷ��ص����á�RNA��ϸ������ֻ����������ͳ�ƺ˺��Ǻ��ᣨnRNA��;���о����������������RNA��ǰ�壬��Щǰ���ڰ��������ɺ���һ���ӹ���������ɸ��ֳ����RNA���ٽ�����ʷ������ã�ֻ�м�С�����Ժ˵�������ʽʼ�մ����ڰ��ˣ�����������ʣ����ǵķ��Ӷ���С����ͳ��С�˺��Ǻ��� (snRNA)���书���Dz�������RNAǰ��ļӹ���������RNA�෴��DNA��Ҫ�ֲ���ϸ�����У�ϸ�����к�������������ϸ������Ҫ��������������Ҷ�����С�
������������RNA��DNA��ȫˮ����ɲ��������������ղ����һ��Ϊ��़����ʵ����������һ�㶼�Ծ��ԣ���ͳ��Ϊ��़�����ʼRNA�����е���़���Ҫ�ǰ����(C)�������(U)��DNA�����е���़���Ҫ��������� (T)�Ͱ���ड�RNA��DNA���������������ʼ���Ҫ����������(A)��������(G)���ڶ������Ϊ���ǣ�RNA�����е�����ΪD-���ǣ�DNA�����е�ΪD-2-�������ǡ����������Ϊ���ᡣ������ˮ��ʱ���������ɶ����м����������ɵ��Ǹ��ֺ����ᣬ���������ˮ����������Ӧ�ĺ��պ����ᣬ�����ٱ�ˮ����������Ӧ�����Ǻ���Ӧ�����ʼ����़ʵ���ϣ������һ���ṹ�������������������Ӱ���һ��������˳��ۺ϶��ɵĶ��������������������ֿ�ͨ������������������ɣ��γ��۵����������еȷ�ʽ�ĸ����ܵĶ����ṹ�������ṹ������ķ��������ܴ�������⣬С�߽������߿ɴ�������������ǧ��������١���������������ʼ��़�϶࣬�ʶ����������������ʣ���������ղ���Ϊ260nm�����ȡ�ǿ�ᡢǿ����صȾ����ƻ���������е������ʹ֮���ԡ����Ժ��������ص�������������������ǿ����������ν��ɫЧӦ���������Ժ�ճ�Ⱦ����ͺ�������Է����ı��ɥʧ��
���������ѧ����ֱ��40����ſ�ʼ�����˽⡣�ر���60�������ȫȷ����������Ŵ���Ϣ�������������ʻ���������֮��������Ŵ���Ϣ��Ԣ��DNA���ӵ�һ���ṹ֮��;���Ŵ���Ϣ�Ĵ���������DNA�����ĸ��ƣ����ﻹ��mRNA��ý�飬����tRNA��rRNA�IJ��롣���ϸ��������˿������ԭ��ϸ�����м����Զ�����ʱ��ͨ�� DNA���������ƿɰ��״��������Ŵ���Ϣ�����ش����Ӵ����Ӵ�DNA��ϸ���˻���������п���Ϊ�ϳ�RNA��ģ�壬���������״���õ��Ŵ���Ϣת¼�������mRNA����mRNA������ʺ��ֿ���Ϊ�ϳɵ����ʵ�ģ�壬������tRNA��rRNA���������ӵ�Э���£������Ŵ���Ϣ���������Ľṹ�����ʻ��������칦�ܵ�ø��ijЩ���ػ���ȹ��ܵ����ʣ��Ӷ������˸�������������ʴ�л��ʽ���ֻ������������������������Ŵ���״���Ŵ���Ϣ���˹��ɽ��д��ݺͱ���Ĺ��̣�һ���Ϊ���������Ŵ������ķ��������㷺�����ڴ��������������������RNAҲ��ֱ����Ϊ�Ŵ���Ϣ�������ߣ�Ҳ�ܽ��и��ƣ�����RNA����������ˡ���Щ������RNA�ڼ���ϸ���м��Ǹ����Լ��Ļ������Ǻϳ������������ʵ�ģ�塣���⣬������һ����ʹ�������°��� RNA���������ǵ�RNA�ڼ���ϸ���п�����Ϊģ�壬������ת¼ø�Ĵ����ܺϳ�һ���µ�DNA������Ϊ��RNA�����ڼ���ϸ����������ֳ�������ߡ��ɴ˿ɼ����������Ŵ���Ϣ���Ǿ��Զ�����DNA����RNA����ЩҲ�ɴ�RNA����DNA�����Ϊ��ת¼���á���˾�������ķ����ѱ�������������ͼ1��������RNAΪģ����ºϳɵ�DNA�����к���ԭ�������°����������������ϵ�����ϸ���Ļ������ڲ�DZ�����У���ϸ�����Ѷ�����ഫ������ijЩ����£�DZ���IJ����°�����ɱ�����Ӷ����ܵ�������ϸ���İ��䡣
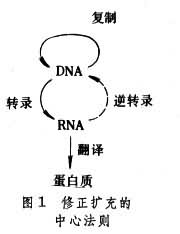 �������Ǽ�����(DNA)
�������Ǽ�����(DNA)
������Դ��DNA����ֻ��������(A)��������(G)�������(C)���������(T)�����ֻ������������DNA�к�����5-�������(m5C)��5-�Ǽ������(om5C)��ϡ�м��������DNA�ļ����ɱ����ձ���������й��ɺ��ص㣺
��1��A��T������ȣ�G��C������ȡ�
��2���������������ԡ�
��3������֯���������ԡ�
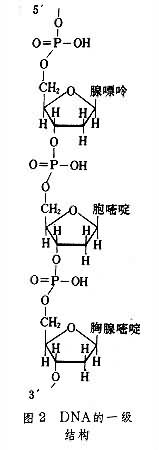
��4����������������䡢Ӫ�������������������Ӱ�졣����������� D-2�������Ǽ������γ���Ӧ�����������ᣬ��Ҫ������������(dAMP)������������(dGMP)������������(dCMP)������������(dTMP)�������ǹ��� DNA����һ���ṹ�Ļ�����λ��ÿ���� DNA�ɺ����������ǧ����˵Ⱥ����ᣬ����֮��ͨ��3'��5'-����������˴��������γɳ����Ķ�����������˼�DNA��һ���ṹ��ͼ2����
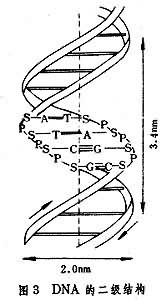
����DNAһ���ṹ�ĺ��������еIJⶨ������70������ڲű�A.M.����ɣ��W.���������Լ�F.ɣ�����ͻ�ƣ��� DNA�Ķ����ṹȴԶ��1953�꼴��J.D.��ɭ��F.H.C.�������ȷ��������DNA�����ɵ��ص��Լ�X��������ķ������������DNA�Ķ����ṹΪ˫�����ṹ�����ѱ����֤ʵ���˽ṹ֮�ص㣨ͼ3�����£�
��1��DNA�������������������������ɣ�������������ʽ����ͬһ���������������������������෴�������ʹ�������γ���ƽ�����е�˫������
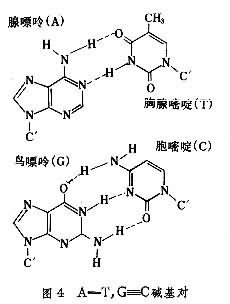
��2���������еļ����λ��˫�������ڲ࣬����֮���������������γɼ���ԣ��Ӷ�ʹ�������ȹ̵ز���������ʵ��������������G��C��A��T֮������������γ�G��C��A=T����ԣ�ǰ�ߺ�������������ߺ�����(ͼ4)��
��3��ÿ��������е��������������ͬһƽ�棬��ƽ����˫������������ɴ�ֱ������ƽ�������±˴�ƽ���ص���ƽ��֮��ľ��붼Ϊ0.34nm������֮�仹�������ŷ����¡��߶�˹������������ˮ����ʹ˫���������ȶ���
��4��������ֱ��Ϊ2nm��ÿתһȦ������Ϊ3.4nm ��ÿ�������ڶ���10������ԡ���Һ�����ϸ���е� DNA������������ʽ��˫�����ṹ�����ڣ�ͨ��B��DNA����B��DNA����������ʪ��С��70��ʱ�����ת��ΪA�͡�A��DNA�ļ����ƽ�治��ֱ��˫�����������ᣬ���������γ�20����б�������Ϊ2.8nm��ÿ����������11������ԡ�A��B���Ϳ��Ի��䣬����DNA��RNA�ӽ�ʱ��B������ת���A�͡�ijЩ����DNA��ϸ��DNA�Լ�������DNA�ȣ���˫���������ṹ�����γɻ�״��Ť�����黨״�����������ṹ��
DNA�Ǿ������������Ļ�����ɳɷ֡�����������У�DNA��Ҫ�ֲ���ϸ�����У�Լռϸ����DNA��98���������Լ�����ļ����鵰���������ԡ����鵰��Ⱦɫ�������ʡ���NHCP����ͬ����Ⱦɫ�ʡ��ݵ���������X�������������Ⱦɫ�������������С�嵥λ����������ɵĴ�������˿�ṹ��ÿ����С�嵥λ�ɺ��Ŀ�������������������ɡ�140������Ե� DNAƬ����������8�����鵰���γɵ����ΰ˾����ϣ����ɺ��Ŀ��������Ƶ�Ȧ��ԼΪ��Ȧ���������Ŀ���֮���Լ��ʮ��������ʮ������Ե�DNAƬ���������˼��������������������ͻ��Ⱦɫ�� DNA��Я���������������ȫ���Ŵ���Ϣ�����ʣ����������������н���Щ��Ϣ������һ����ϸ���ʵ��������Ҷ������Ҳ����������DNA��Լռ������2��������DNA�ķ���������С�����뵰���ʽ�϶����ɴ��ڣ����γɻ�״��ֻЯ������ϸ����������ijЩ�����ʵ��Ŵ���Ϣ��Ҳ�ܽ��и��ƣ�����Ϣ�����´����ʳ�����Ϊ����Ⱦɫ�塣
DNA�ĸ�����������ϳ����Ŵ����ʵ�һ�ָ��ӹ��̣��˹��̴������£��������ض�������ʶ���DNA�����ϵĸ���ʼ�㣻�ڴ���ʼ�㣬�������칹ø������ø��˫�������һС��;��RNA�ۺ�ø�ϳ�һ��RNAƬ�Σ���Ϊ�ϳ���DNA��������Խ��DNA�������ֱ�Ϊģ�壬�� DNA�ۺ�ø�������£���������������������(dATP��dTTP��dGTP��dCTP)Ϊԭ�ϣ���5'��3'����ֱ���кϳ�����ģ��Ļ�����������һ��Ϊ�����ϳɵ�����������ǰ���ɣ���һ��Ϊ�����ϳɵ�����������Ƹ���Ƭ�Σ��� DNA����ø�Ĵ��£��������Ƭ��ƴ�������������ӻ��ɡ��ӻ��ɵij�����ǰ������ȣ��������෴����˿�����DNA�ڸ��ƹ����п���һ�����״�DNA������������ȫ��ͬ���Ӵ�DNA������ÿ�����Ӵ�DNA�ж��������״�DNA��һ��������DNA�����ָ��Ƴ�Ϊ�뱣�����ơ�DNA�İ뱣�������������Ŵ������Եĸ��ݣ�����֤���������ֵ��ȶ����������ԡ�ͨ�� DNA�ĸ�����ʵ���Ŵ���Ϣ��ת�ƣ�һ������ͬһϸ���ڽ��еġ�����Щ����£�ijЩ�Ŵ���ϢҲ�ɴ�һ��ϸ��ת�Ƶ���һ��ϸ����ȥ����ʹǰ�ߵ�ijЩ�Ŵ���������״�����ں���֮�С����ֲ�ͬϸ��֮���Ŵ���Ϣ��ת�ƣ�������Ȼ��ϸ��ת����ת�������Լ��˹��Ļ��̣�ʵ���϶��Dz�ͬϸ��������DNA֮������顣
DNA�ڸ��ƻ����������������ֲ������DNA���ӱ����ܵ����ijЩ�������ص����ã�����ӽṹ���ɸı䣬�ر������еļ���ɷ������иı���ƻ�����������£���������һ���̶��ڽ������;�������֤�Ŵ���Ϣ�ܹ���ʵ�ش����ഫ������� DNA�ĸı�������ϵ��ȱ�ݶ����ܻ�δ����������������ͻ����������ͻ����ʽ��Ҫ�У�
��1��DNA�����м������ת�������ǰ����ָ DNA�еļ���������ʼ�֮�䣨A��G������़�֮�䣨T��C���Ļ��䣬������ָ���ʼ����़�֮�䣨����A��T��G��C���˴˵Ļ��䡣
��2��DNA�����з������룬��DNA�ļ�������мӽ���ʧһ�������������������ӵĻ��ҡ�ǰ�ƻ���ơ�
��3������ DNA�ۺ�ø��������ظ���ϣ����Ƶ��Ӵ�DNA�����ж�ʧһ�λ�һ��DNA����
��4������IJ����������������������������ʹDNAһ���������ڵ�������़��Թ��ۼ�����������������Щͻ����ʽ���ɵ��� DNA�ĸ����Լ�ת¼�ͷ�����ϰ�����Σ�ʹ������������졢������������������������������ϸ��ƶѪ֢��һ�ֵ��͵ķ��Ӳ�������DNA������Ѫ�쵰�����һ�����е�CTT�����ӱ����CAT�����ӣ�ʹѪ�쵰���Ӧ����е�6λ�ĹȰ��������Ӱ��ᣬʹѪ�쵰�����ʸı䣬��������ɥʧ��
ת�˺��Ǻ���(tRNA)tRNA�㲼�ڸ���ϸ���İ�Һ��λ������Լռϸ����RNA��15����������״̬���백�����ϳɰ�����tRNA����ʽ�����ڡ��������ᴿ��ȷ���˺��������е�tRNA������ 200���֡��Ƿ�������С��RNA��������ԼΪ2.4��3.1������٣���������Ϊ4S������ǰ�ֳ��� 4SRNA��tRNA�Ĺ������ڵ���������ϳ��н�������Ӱ�Һת�˵���˵�������ȥ��һ��tRNAֻ��ת��һ������İ����ᣬ��һ�ְ�����ɱ�����tRNAת�ˣ�������tRNA��Ϊͬ��tRNA��
tRNA���ӵ���ɵ�λ��Ҫ�ǣ������ᣨһ��������AMP����������(һ��������GMP)�������ᣨһ�������CMP���������ᣨһ��������UMP���Լ��������������ζ�����ϡ�к����ᡣÿ����tRNA��74��94���˵Ⱥ����ᰴ��һ��������˳��ۺϳ�һ�����������������������֮�䶼ͨ�� 3'��5'-����������˴�������ͬ��tRNA�ĺ��������ж��dz����ƣ������ַ�ͬ��tRNA�ĺ���������֮������ںܴ�IJ��죬����ʾ�ṹ�빦����һ�������һ���ԡ�
����tRNA�Ķ���������������۵����γɹ�ͬ��ʽ����Ҷ���ζ����ṹ(ͼ5�����˽ṹ��������۵�����ͨ��������ۡ�����������ۡ���������ऱ�(DHU��D��)���������ӱۣ�AC�ۣ����ɱ��(�����)�����ռ����հ���(T��C)�ۡ�ÿ�������ɡ������͡��Ǿ�����������ɣ������еļ�������������ɼ�������ԣ����γ�һ��С��˫�������Ǿ���λ�еļ���������������ɼ���ԣ������Ե�����ʽ���ڣ����γ�һ��С�����뾥��������������ν����ʽ�ṹ����ת�˰�����ʱ�������۵ķǾ���λ�����ж��صĹ��ܡ����磺
��1������tRNA�İ�����۷Ǿ���λ���������3'���ǻ������ض���������Ȼ����ϳ�������������Ӧ�İ�����tRNA������������ʽ��������Ӱ�Һ������˵����塣
��2���������ӱ��еķǾ���λ���������ӻ��������ض��ķ������ӣ���ʶ��mRNA����Ӧ�������ӣ�����֮������ϣ������ɱ�֤mRNA���ض����Ŵ���Ϣ���������ĵ����ʡ�
��3��T��C�۵ķǾ���λ(T��C��)����˵������5SRNA��5.8SRNA������ϣ���ʵ��tRNA��˵�������ǻ��Ľ�ϣ����tRNAת�˰����������tRNA����Ҷ���ζ����ṹ���ɽ�һ�����ɸ��ȶ��������ṹ��
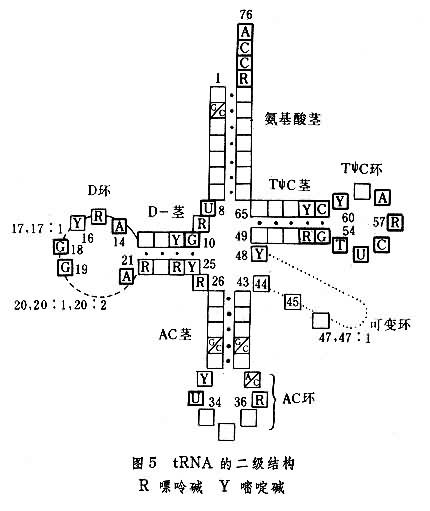
tRNA����һ����ͬ���ص㣬������������ϡ�к���(��)�϶࣬�����ɴ�5��20���������70���֣����Ƕ�����tRNAȫ����ˮƽ����ǰ��ˮƽ���ɻ�������(��)���ζ��������ֳ����κ��ա�������������Ҫ��ͨ�������Լ�����ԭ���Ѱ�������ӵķ�Ӧ�����β�λ�ɷ����ڻ������յļ���ϣ������ں����ϣ�Ҳ����ʱ�����ڶ���֮�ϣ����⣬���ɷ����ڼ���ͺ��ǵ����ӷ�ʽ�ϡ��������tRNA�����ζ�һ�����ԭ��������κ��մ��λ��tRNA�����е��ض���λ��������֪����tRNA�ķ�������3'���Աߵĺ����ᶼ��ϡ�����ʺ����ᣬ���Ƕ��ϱ𡢼ල���ٽ�mRNA��������tRNA�������ӵ�ȷ��Ͼ�����Ҫ���á�
��ʹ���Ǻ���(mRNA)�������ں�mRNA���٣�Լռ��RNA��5�����£�������״̬�������˵������ϳɴ������Ķ�˵��������ʽ�����ڰ����С�ϸ����mRNA������࣬�����Ӵ�С������⣬һ�������ٸ�����ǧ����������ɣ�������ԼΪ20��200��������ԼΪ6��30S��mRNAһ�㶼���ȶ�����л��Ծ������Ѹ�٣������϶̣�ԭ��mRNA�İ�����һ��ֻ�������ӣ������һ��Ҳ������Сʱ�����졣mRNA���ӵĻ�����ɵ�λҲ��AMP��GMP��CMP��UMP��������ϡ�к��գ�ԭ��mRNA���������٣����mRNA������Щ�����ᰴ��һ��������˳��ͨ��3'��5'-����������˴��������γ�һ���ܳ��Ķ�������������������жദ�۵��ɷ���ʽ�ṹ�����γɶ����˫����������ͬ��mRNA��������ʽ�ṹ����Ŀ����ɡ������Լ�����λ�õȾ�������ͬ�������ǵĶ����ṹû�й�ͬ��ģʽ���ɡ�
ԭ������mRNAһ�㶼�Ƕ�˳��������ʹ����һ����mRNA���м��ֵ����ʵ��Ŵ���Ϣ������Ϊ�ϳɼ��ֵ����ʵ�ģ�塣�෴���������mRNAһ�㶼�ǵ�˳��������ʹ����һ����mRNAֻЯ��һ�ֵ����ʵ���Ϣ��ֻ����Ϊһ�ֵ����ʵ�ģ�塣���mRNA������ת¼���ɵ�ǰ��(hnRNA)���ӹ�������������ԭ��mRNA����������ǰ�壬���DZ�ֱ��ת¼���ɱ�ֱ�ӽ��з��롣
mRNA������������ģ�����õIJ�λ��Ϊ��������������λ��Ϊ�Ƿ��������������в��ֺ��������о��б������ã����Ƿ���������ȫû�б������á����mRNAֻ��һ���������������Ƿ�������ԭ��mRNA��ɸ����������������д� DNAת¼�����Ŵ���Ϣ��������������˳�������֣�����˵�������������ÿ3�����������һ�������壬����Ϊһ����Ϣ��λ��ͨ��һ�������ӡ�����mRNA����4�ֲ�ͬ�������������л���ɣ���ô�ؿ����г�43=64�ֲ�ͬ��������������ӡ����г�UAA ��UAG��UGA�����⣬����ÿ�������Ӷ�����һ������İ����ᣬ��һ�ְ�����ɱ����֣�1��6�֣��������������������������ӳ�Ϊͬ�������ӻ�������ӡ����ص���ǰ���������ᶼ��ͬ��������������ȫ��ͬ��UAA��UAG��UGA3���������䲻�����κΰ����ᣬ���Ե����ʺϳɵ���ֹ���о��������ã��ʳ���ֹ�����ӡ�AUG�����ǵ�����������ӣ����һ������������õ����źţ��ʳ��������ӡ�GUG���Ӱ���������ӣ�����ԭ��������Ҳ�����������ӡ������Ƿ������䲻�߱����ģ�����ã������ǵ���ʼ��λ������ֹ�����ӣ��ɱ�֤������̰�ʱ��ֹ�����⣬�����Ƿ������ж���һ�θ������ʺ���������У���ԭ��ϸ���˵�����С�ǻ���16SrRNA��3'ĩ�˶���һ�θ�����ऺ���������У���5'��CCUCCU��3'�����߿��Ի�����ϣ���ʵ��mRNA��˵�����Ľ�ϣ��˽�Ϸ�ʽͨ���Ķ��������ŵ���Ϸ����ǵ����ʺϳɱز����ٵ�����֮һ��
���������� RNAһ�������mRNA�ϳɵ�ȫ����Ҳ����ת¼�ͼӹ������Ρ�ת¼�����������ɲ����Ե�mRNAǰ�壬��hnRNA���ӹ��������ǰ�������л��Եij���mRNA��ת¼�����ڰ����н��У������� DNA��ģ�壬���������������(��UMP��CMP��GMP��AMP)��ԭ�ϣ�����DNAָ����RNA�ۺ�ø�Լ�Mg2+��M �Ȳμӣ����������DNAģ�廥����mRNA������ȫ���̰���ʶ����ʼ�����켰��ֹ�ĸ��Ρ����ת¼���ɵij�������ΪmRNAǰ�壬�뾭���м����ӹ��������ܱ�ɳ���mRNA��
�Ȳμӣ����������DNAģ�廥����mRNA������ȫ���̰���ʶ����ʼ�����켰��ֹ�ĸ��Ρ����ת¼���ɵij�������ΪmRNAǰ�壬�뾭���м����ӹ��������ܱ�ɳ���mRNA��
��1���ڶ���ø�Ĵ��£�����5'�˼���һ����7�����յ���ν��ñ�ӡ���5'��ñ������ֹ5'���к���ø�����ã�����mRNA���ȶ����Լ����뵰���ʺϳɵ������á�
��2���ڡ����A�ۺ�ø���Ĵ��£���3'ĩ������һ����ν�����Aβ�͡�������Լ��30��200�������ᡣ
��3����snRNA�����к���ø������ø�İ����£������������ޱ�����Ե��ں����г�����ʹ�����л��Ե�������ƴ��������ʹ֮��Ϊ�����RNA���Ž�����ʷ������á�
�˵�������Ǻ���(rRNA)��ϸ���к�������һ��RNA��Լռ��RNA��80�����ϣ��ڰ������뵰���ʽ�ϳɺ˵��������ʽ�����ڡ�������Դ�ĺ˵����嶼���ɴ�С��ͬ�������ǻ����ɣ�ÿ���ǻ��������ֵ����ʺʹ�������������rRNA��ԭ��ϸ���˵�����Ĵ��ǻ�������rRNA����23SrRNA��5SrRNA���ֱ���3000����120����������ɣ�С�ǻ�ֻ��һ��rRNA����16SrRNAԼ��1500�������ᡣ���ϸ���˵�����Ĵ��ǻ�������rRNA������5SrRNA�⣬����28SrRNA��5.8SrRNA������߷ֱ���5000��150����������ɣ�С�ǻ�Ҳֻ��һ��rRNA����18SrRNA����2000����������ɡ�����rRNA��һ���ṹ��Ҫ������AMP��GMP��CMP��UMP���ֻ�����������ɵ�һ�����������������5SrRNA�⣬��������rRNA������������ϡ�к����ᣬ������Ҫ�Ǽ����Ļ���������ͼ�������(��)���������rRNA���߶���ԭ�����ߡ�����rRNA���Ķ����ṹ��ʽҲ������ͬ��rRNA�Ĺ��ܳ��Ժ˵��������ʽΪ����������ϳ��ṩ���ʵij����⣬5S��5.8SrRNA����tRNA������ϣ�16SrRNA����mRNA������ϣ�������Ƕ������ϳɵ����������ֱ�ӹ�ϵ��
С�˺��Ǻ���(snRNA)���ϸ�������·��ֵ�һ�ֶ��ص�RNA����15��֮�࣬����������RNA��1����ʼ�ղ�������ʣ�����������С��Լ��90��400�����ᣬ������������6�ֶ����������ᣬ�����ɴ�35��������ΪU-snRNA������U1��U2��U7�����������о��Ѹ��壬����������Nm��ϡ�к��գ�ǰ���ּ�U7��5'�˾���һ����ͬ�ṹ�ġ�ñ�ӡ�����ñ����mRNA��5'����Ϊ���ƣ�ֻ������7-��������(m7G)��2��2��7-����������(m����27G)�����棬�����ṹ��ȫ��ͬ��U-snRNA�Ĵ�л�ȶ������Ժ˵�������ʽ���ںͷ������á�����ֻ��U3-snRNA�����ڰ��˵ĺ����У���rRNAǰ��ļӹ��йء��������־������ڰ��˵ĺ����У�����RNA�ۺ�ø���ת¼���������ܿ��ܸ��죬����U1��U2��mRNAǰ��ӹ�����ʱ�ز����ٵ����ء���Uϵ��snRNA��֪����ʮ���֣������ں��ʻ���ʣ�����RNA�ۺ�ø���ת¼���ɣ�5'�˾������κ�ñ�ӣ�����Ҳ���ܸ��졣