[ƴ��]��mianyi qiudanbai
[����]��immunoglobulin��Ig
һ���������Ļ�ѧ�ṹ�����߹��ܵ��ף���������Һ�ں��ܰ�ϸ�����棬�ǿ�������ʻ��������ṹ���ܵIJ�ͬ��Ϊ�����:IgG��IgM��IgA��IgD��IgE������Ӵ�С����ɡ���������ɺ�̼ˮ�����ﺬ���ܲ���һ��������̬��Ӧ�Լ��������������Լ��������ָ�Ⱦ�Լ���Թ�������ʱ���������쳣���ߡ������������ȱ���ۺ���������ȱ��ʱ�������쳣���ͣ��ٴ��ϳ�Ӧ���������Ƽ����δ�Ⱦ����ijЩ����������ȱ�ȡ�
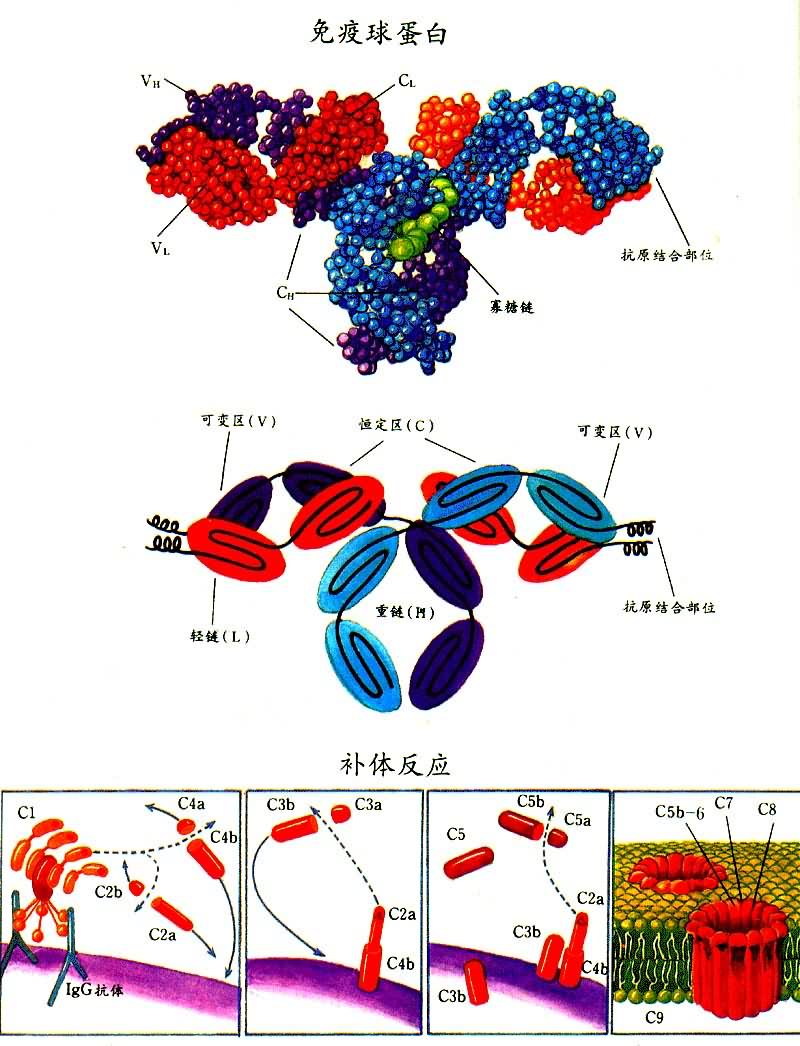
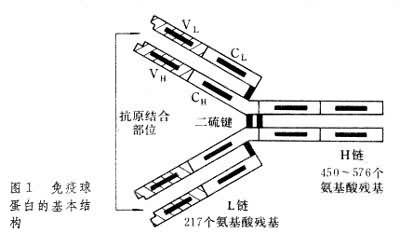
�����Ļ����ṹIg���ӵĻ����ṹ�ǶԳƵ���������������������ͬ��������L������������ͬ��������H����������������������ɡ�Y�����͵�����ӣ�ͼ1����Ig���Ӳ�ͬ������ɸ��������Ľṹ�����ġ�������Ϊ�á��̡��ġ��ļ��������࣬�ֿɰ�������ԭ���IJ�ͬ��Ϊ�������ࡣ����Ig���ӵ���������ͬ���ɷ�Ϊ���ͺͦ��ͣ����ͼ�İ�����Ϳ�ԭ�Բ�����ͬ����������������ĩ�˵İ�����˳�������仯�����ⲿ�ֳ�Ϊ�ɱ�����V������V��������ʶ��ԭ�;������������ԵIJ�λ���������ಿ��(���Ȼ�ĩ��)���������Ŀ��˳����Ժ㶨���ʳ��ȶ�����C�������ȶ���������Ig���ӵĸ������﹦�ܡ�Ig����Ҫ���ﻯѧ���ʼ���1��
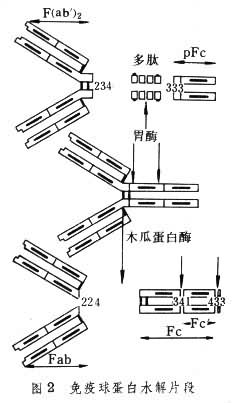
��ľ��ø�ɽ���������Ϊ������С���ƵĶ�Ƭ������������Ƭ���뿹ԭ��ϣ���Ϊ��ԭ���Ƭ��(Fab��)����һ��Ƭ�ɽᾧ����Ϊ�ᾧƬ��(Fc��)����θø�ɽ���������Ϊ���Σ�һ�����ƬΪ5S˫��F(ab��)2��һ��С��Ƭ��Fc���ƣ���ΪpFc�䣨ͼ2����
 ������(Ig)���ӵ�����
������(Ig)���ӵ�����
Ig �����������Ŵ����ƵĿ�ԭ�Բ�ͬ������������ԭ��������ͬ�����в�ͬ�ı����ԡ�������
��1��ͬ�������ԡ�ͬһ���������г�Ա��Ig���Ӷ��߱��Ŀ�ԭ��ǡ����磬��������������������ף�����IgG�ɰ�����������IJ�ͬ��ΪIgG1��IgG4�ĸ����ࣻIgM��IgM1��IgM2�������࣬IgA��IgA1��IgA2�������࣬ IgD�� IgD1��IgD2��������ȡ�
��2�����������ԡ�ͬһ������ͬ��Ա��Ig���ӿɾ��в���ȫ��ͬ�����쿹ԭ��ǡ�
��3��������(Id)��ÿһIg���ӽԿɾ��ж��صĿ�ԭ�����ء��������ھ���ǧ����������γ�ϸ����¡��ϸ���꣩���������������Ϊ�����ࡣ
�������ӵ����﹦��Ig���Ӿ��н�Ͽ�ԭ�ʹ̼��������ɵ�˫�ع��ܣ���2�������ȣ������뿹ԭ��ϣ�������������ЧӦ��������
��1���벡ԭ�����������ڵĶ��ؽ�ϣ���������Ⱦ���ߣ�
��2�����Һ��һ��������֣���������ӣ���ɱ�˲�ԭ����ϸ�������ã�
��3����ǿ����ϸ��������ϸ�������ɻ�ɱ��ЧӦ��
��4������֯�еķʴ�ϸ�����ȼ�����ϸ����ϣ�����������Ӧ��
��5�������ֲ����������ǿ�����ı����������ų⣻
��6���������ϸ�����������߱������������ܴ���̥������̥�������⣬����Ig�������ǵ�����ɣ����Գ�������������ԣ����п�ԭ�ԣ��ɻ��������ϸ����ʹ֮������Կ���Ŀ��婤���������Ϳ��壨Id���壩���Ӷ��γ��������ڵĹ��ܡ�
 ��������������
��������������
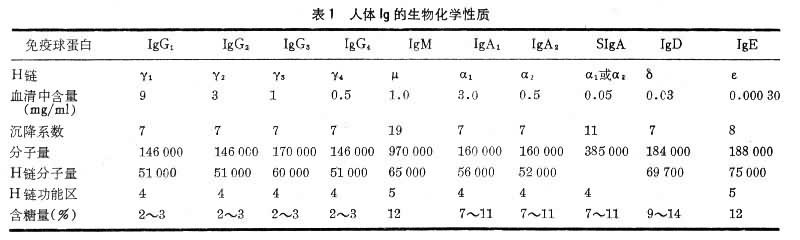
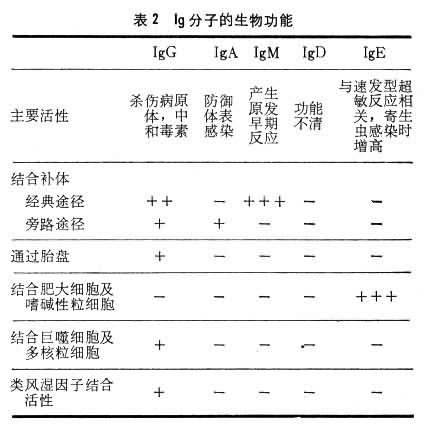
����Ig������������ѧ�����ϸ��в�ͬ��
��1��IgG��IgG��������Һ����Ҫ��Ig��ԼռѪҺ��Ig������70��75��������IgG��ͨ��̥�̣�������������ĸ���õ� IgG�ڵֿ���Ⱦ��������Ҫ���á�Ӥ��������2��4�ܿ�ʼ�ϳ�IgG��8���Ժ�Ѫ����IgG�ɴﵽ����ˮƽ������IgG��������Ig������ɢ��Ѫ����ļ�϶�ڣ�����ڽ�ϲ��塢��ǿ����ϸ�����ɲ�ԭ������к�ϸ�����ص��������棬������Ҫ���ã�����Ч�ؿ���Ⱦ�����Ƕ�����������һ�档��ijЩ�������߲�����������������Ѫ��ƶѪ��ѪС�������������Ǵ��Լ����ʪ���е��������嶼��IgG��һ��������Ӧ������ϸ����ϣ������ǿ����֯�������á�
��2��IgM��IgN��Ig�з��������ͨ����Ϊ���ף�ռѪ��Ig������10���������������¹۲죬IgM����������ṹ��ͬ�ĵ�����ɡ�����λ����һ��������(J��)����ɡ���״��������塣IgM���ڸ��巢����������������Ŀ��壬Ҳ�Ǿ���ԭ�̼��Ķ����������ȳ��ֵĿ��壬��˼��IgM�ĺ����������ڴ�Ⱦ����������ϡ�IgM��̥��3���º�ʼ�ϳɣ���ˮƽ�ܵͣ�1��2��ʱѪ����IgM�����ﵽ����ˮƽ��ͨ����ϲ��壬IgM���ܽ�ϸ�����ܽ�Ѫϸ�������ã������кͲ�������Ч�ܱ�IgG��100�����ϡ��ܶ�������Ȼ������ͬ��Ѫ���أ���A���뿹B��Ѫ�������ʪ���е����ʪ�����Լ�÷���IJ����Ͽ��嶼����IgM��
��3��IgA��IgA��Ѫ���еĺ���������IgG��ռѪ��Ig������10��20����IgA�е��壨1�������ṹ����˫�壨2�������ṹ�������壨���ɸ������ṹ����J�����ᣩ�Ȳ�ͬ��ʽ��Ѫ���е�ΪѪ����IgA����ҪΪ7S���塣���ַ���Һ������Һ�����ᡢ��Һ�����顢������������������Һ�е�IgAΪ������IgA(SIgA)���ɶ����弰����幹�ɣ�����з���С����ڡ�����С�������ڷ�����IgA�ֿ�����ø��ˮ��ʹ�ʹIgAͨ��������֯��ճĤ�������Һ�ڡ�������IgA�������Եı��������������ԭ���ֵĹ��ܡ�
��4��IgD��IgD��Ѫ���ں����ܵͣ�����������1������IgD��IgG1��IgG2��IgA��IgM���ױ�����ˮ��øˮ�⣬���������ܡ�IgD �����﹦���в�ʮ���˽⡣Ŀǰ��֪��IgD�����������ϸ���˿��塢������Ĥ���塢���ȵ��ؿ��塢���������Ѫ�� O���塢����ù������Ϳ������ص���������IgD�뼲���Ĺ�ϵ���˽ⲻ�ࡣ
��5��IgE������Ѫ���� IgE�������͡�IgE��Ҫ�ɺ������ͳ����ܰͽ��еĽ�ϸ���ϳɡ��ڱ�ǻ��֧���ܷ���Һ����֭����Һ�д��ڷ�����IgE��IgE��һ����ϸ�����壬����ѪҺ�е��ȼ�����ϸ������֯�еķʴ�ϸ���Լ�Ѫ����Ƥϸ����ϣ��������۵ȸ��ֹ���ԭ����ԭ��IgE����Щϸ�������ϣ�ʹ֮�ͷŴ������Խ��ʣ����鰷�ȣ�����շ�I�ͱ�̬��Ӧ��
��������������ܲ���ǧ�����ֲ�ͬ���������ӣ��������н������ڵĿ�ԭ�����������Ӧ�Ŀ����������Է�Ӧ���ܹ�������˴���������Ŵ����ʣ������Ӻζ�������Ҫ������ѧ˵�Դ˼��Խ��͡�
��1����ϵѧ˵�����ڹ۵���Ϊ���������������䱸��ȫ�����ڱ��뿹����ӵ�ȫ��������ͨ����ֳϸ���Ŵ���ȥ������֪��ÿ���ܰ�ϸ���Ծ���Ig���ӵ��Ŵ���Ϣ��������VL��VH��������ṩ������ֿ�ԭ��ϵ�λ�㣬����Ig������״����Ӵ���ֱ�Ŵ��Ĺ�ϵ����֤���Ӵ�ϸ��������V��D��JƬ�εĶ����ԡ�
��2����ϸ��ͻ��ѧ˵����Ϊÿһ����ֳϸ�����Ǿ����γ�Ig���ӵ�ȫ���Ŵ���Ϣ�������̳�������V�������ڸ��巢�������У�������ϸ��ͻ�䣬V������������ԡ�Ϊ�ˣ�ÿһ����ϸ���Կɱ��ﲻͬ�������ԡ�
��3���������ѧ˵��Ig�ɱ���(V��)��Ϊ VƬ�Ρ�DƬ�μ�JƬ�Ρ���Щ����������ϣ����ɾ���һ��V-J����������;���һ�� V-D-J�������������������ϳɺܶ�����ᡢ������ V����V���������벻ͬ��C��������������������Ļ����������Ļ����ֿ�������ϳɸ����Ig����
��4�����Ӷ��������ۡ�����������ĵ�3�ɱ����е�96λ������ĺ����ᣬ����V��JƬ�ν��ʱ˳��IJ�ͬ�����������ԡ���������˳��Ķ�����Ҳ�����������С�
��5��������D������Ķ����ԡ�D�������Ig������ʱ���������ֿ�ܽ��з��롣����������ʵĿ�ܣ����Ϊ��ǰ��ܻ�����ܡ�ÿһ�ַ�����ʽ����ָ��һ��DƬ�η���ɶ��ְ������˳���嵽һ��Bϸ��������ȡ��һ����ʽ����ȡ���ڲ�������������ԡ�
��6��С����������ۡ���DƬ�κ�JƬ����������У�V������Կ������С���������߶ȿɱ�����
������˵����һ�������£���ԭ����Bϸ����ϸ���γɣ���VDJ�����£����ں��в�ͬ���������У���ͬ�ռ乹�͵�Ig���ӡ������ǧ���Ŀ�ԭ��������Բ�����Ӧ�ľ��в�ͬ��ԭ��ϲ�λ�Ŀ��壬�������ֶ���������Ӧ��