[拼音]:shengtai celüe
[外文]:ecological strategy
生物在进化过程中形成的对付环境的策略。这里的环境不仅是物理环境,而且还包括同种和异种的其他生物。因此,生态策略也就是生物在广义的生存环境中形成的进化策略。
近代研究进化论的一些学者常假定自然选择是一个最优化过程,认为现存物种代表了在各种可能的适应方式中的最佳选择,因此常采用工程学家和经济学家提出的最优化理论来研究生物进化。在近代生态学文献中,生态策略的概念被广泛接受并得到迅速普及,出现了生殖策略、防卫策略、资源获取和分配策略、取食策略、生活史策略等等概念。随着研究的深入,人们注意到,某一方面(如防御)的最佳策略不见得是另一方面(如取食)的最佳策略。因此人们又开始将上述种种策略综合起来进行整体研究,探讨生物进化的总体最佳策略。
1954年,英国鸟类学家D.拉克在研究鸟类生殖率进化问题时提出,生殖率和动物其他特征一样,是自然选择的结果;每一种鸟的产卵数表现出一种倾向,似乎以保证幼鸟存活率达于最大值为“目标”。生态学家在自然界普遍看到:成体大小相似的物种,如果产大卵,则产卵数少,如果产小卵,则产卵数多。生物可利用的能量资源是有限的,如果把有限的能量用于保护幼体的存活,则必然难以同时提高生育力。这涉及进化过程中的能量分配问题。也就是说,在自然选择中,动物总是面临着两种相反的、可供选择的进化策略:一种是低生育力,但亲体有完善的照顾和培育幼体的行为;另一种是高生育力,却缺乏亲体照顾行为。
1967年,R.H.麦克阿瑟和E.O.威尔逊推进了这个思想,首次按栖息环境和进化策略把生物分成r策略者和K策略者两大类。在此使用的r和K是与种群增长方程中的内禀增长率r和环境负载量K两个参数具有同样的含义。他们认为,地球表面环境是连续变化的,一个极端是气候稳定、天灾稀少的环境(如热带雨林),属于生态上饱和的系统,动物密度很高,竞争激烈;另一个极端是气候不稳定、天灾频繁的环境(如寒带或干旱地区),属于生态上不饱和的系统,密度影响可忽略,缺乏竞争。在前一类环境中,动物种群数量达到或接近环境负载量水平,即与种群逻辑斯谛增长模型的饱和密度K接近,因此这种环境对生物的作用可称为K 选择。在后一类环境中,种群密度处于K值以下的增长段,因此种群常处于扩展增大过程,而呈现高增长率r,所以这种环境对生物的作用可称为 r选择。麦克阿瑟和威尔逊最初之所以提出r或K策略思想是用来解释岛屿定居现象的(见岛屿生态)。
1970年,E.R.皮安卡把这个思想表达得更详细深入,并扩展到应用于所有生物。他列表比较了r选择和K选择的有关特征(表1)。

在生物大类群间作策略比较,可以把昆虫看成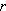 策略者,而脊椎动物是K策略者。脊椎动物主要盛发期是侏罗纪、下白垩纪及始新世和渐新世,正是温暖潮湿气候稳定的地质期;相反,昆虫的快速进化则是在二叠纪和三叠纪,当时的气候条件被证明是非常多变的。在大类群之内,同样可进行策略比较。例如,鸟类中的鹫、鹰、信天翁等是典型的K策略者,它们的体型大,生育力低,对幼鸟有良好保护;而小型的山雀、虎皮鹦鹉等是典型的r策略者。由此可见,r策略者和K策略者是两个进化方向不同的类型,其间有各种过渡,有的更接近于r策略者,有的更接近于K策略者;也就是说,从极端的r策略者到极端的K策略者之间有一个连续的谱。
策略者,而脊椎动物是K策略者。脊椎动物主要盛发期是侏罗纪、下白垩纪及始新世和渐新世,正是温暖潮湿气候稳定的地质期;相反,昆虫的快速进化则是在二叠纪和三叠纪,当时的气候条件被证明是非常多变的。在大类群之内,同样可进行策略比较。例如,鸟类中的鹫、鹰、信天翁等是典型的K策略者,它们的体型大,生育力低,对幼鸟有良好保护;而小型的山雀、虎皮鹦鹉等是典型的r策略者。由此可见,r策略者和K策略者是两个进化方向不同的类型,其间有各种过渡,有的更接近于r策略者,有的更接近于K策略者;也就是说,从极端的r策略者到极端的K策略者之间有一个连续的谱。
近来r-K 策略的概念已被广泛地用于说明杂草、害虫和拟寄生物等的进化策略。在农业生态系统中,人类种植农田并进行施肥和喷药等经营活动,杂草或害虫要生存其中,就必须具有较高的生殖潜能和扩散能力。因此,杂草和害虫(如狗尾草、马唐、飞蓬和豚草等杂草,以及褐飞虱、黏虫、螟虫等害虫),一般都是r策略者。飞蝗可被视为具有两种策略交替的特殊类型:群居相是r策略的,散居相是K策略的。蚜虫的有翅和无翅世代交替也是这样。至于选择拟寄生物作为防治害虫的天敌,同样要考虑r或K策略者对天敌的不同反应。
r和K两类生态策略,在进化过程中各有其优缺点。K策略者的种群接近K值但不超过,因为超过K值有导致生境退化的可能。生育力低则要求有高存活率,这才能保证种族的延续,因此K策略者防御和保护幼代的能力较强。由于有亲代照顾培育,K策略者通常寿命较长,个体较大,这些特征可保证K 策略者在激烈的生存竞争中取得胜利。但是,当K策略者种群在过度死亡或激烈动乱之后,回到平衡水平的能力是有限的。如果种群确实很小,还有可能灭绝。大熊猫、虎、豹等珍贵稀有动物就属此类。因此K策略者的资源保护比r策略者更困难、更重要。
相反,r策略者的密度是经常激烈变动的,常常突然暴增或猛烈下降。因为r≌lnR0/T(其中R0为净生殖率;T为平均世代时间),所以高r值是通过提高生殖率和缩短世代时间达到的。但其死亡率很高,防御和竞争能力不强。高 r值必然导致种群的不稳定性。但种群不稳定并不意味在进化中必然不利,其数量很低时r策略者不象K策略者那样易于灭绝,反可通过迅速增殖而恢复到较高水平。r策略者当种群密度很高时,可能大量消耗资源使生境破坏,但它们通常具有较大的扩散和迁徙能力,可以离开恶化了的生境,在别的地方建立新种群。所以,r策略者的个别种群虽然易于灭绝,但物种整体却是富有恢复力的。如果说,K策略者在生存竞争中是以“质”取胜,则r策略者可视为以“量”取胜,所以有的学者将r策略者称为“机会主义者”,一遇良好机会就会出现大发生。r策略者的死亡率高,扩散力强以及它们需要不断面临新局面,这些特征可能使它们成为物种形成的丰富源泉。
1974年,T.R.E.索思伍德总结了r或K两种策略者种群动态特征间的区别(见图)。

图中对角线代表Nt+1与Nt相等,种群平衡。K策略者曲线与对角线有两个交点 X和S。X是不稳定的平衡点,可称为灭绝点,如果种群下降到X点以下就有灭绝的危险。相反,r策略者由于低密度下增殖快,所以只有一个平衡点S,种群易在S上下作激烈波动。此模式还说明,天敌对于这两个极端的策略者的作用都不大。对于r策略者,由于它增殖比天敌迅速,故其数量不易被控制;待天敌种群发挥作用时,它们已迁出原地,在新的地方形成种群。对于K策略者,由于其个体大,竞争力强,天敌的作用也难以发挥。当然多数动物处于这两极端的中间,天敌的作用仍是重要的。
1977年,J.P.格里姆将植物的生长和生殖策略划分为3类:竞争型、逆境耐受型和杂草型,这3种类型是针对环境中的逆境(指营养缺乏及光照、气温不足等)和破坏(指动物取食、细菌致病或人类损害等)的程度而划分的(表2)。因为有许多试验证明,能在逆境(如严重干旱)中生活良好的植物,不一定是优良的竞争者,因此可以把逆境耐受型和竞争型分别作为独立的型。如果说,杂草型与r策略者相当,则逆境耐受型就可与K策略者相比(因为环境破坏程度低、种群相对较大)。但还要增加一个维度来描述竞争型,有些学者采用了下面的三角形图解。竞争型策略出现在逆境和破坏程度都很低的环境,植物生产量高,种群波动小。有的学者把此种策略称为α策略。α策略的命名是根据 A.J.洛特卡-V.沃尔泰拉二氏竞争数学模型中的竞争系数α 而起的。植物中的异株克生现象和动物间通过干扰作用阻止竞争对手利用有限资源等现象,都是α策略的例子。